I linfociti rappresentano una popolazione eterogenea di elementi leucocitari
distribuiti in varie zone dell'organismo. Nell'accezione classica essi risultano
costituiti da piccoli elementi dotati di scarso citoplasma, con nucleo a
cromatina pienotica; tuttavia attualmente la dizione di «cellule linfocitiche»
tende a far riferimento all'intero complesso di elementi che coinvolge i
linfociti tradizionali (piccoli, medi e grandi), i linfoblasti e le
plasmacellule come forme di attivazione di una definita categoria degli stessi
elementi.
La maggior parte dei linfociti nel sangue periferico è costituita da piccoli
elementi di 7-8 micron di diametro (piccoli linfociti), mentre una quota minore
(10%) è rappresentata da elementi di 9-15 micron (medi e grandi linfociti).
I piccoli linfociti appaiono costituiti da un nucleo con cromatina a zolle
condensate, rotondeggiante o appena indentato, privo di nucleoli visibili,
occupante tutta la cellula ad eccezione di un piccolo lembo di citoplasma
moderatamente basofilo; questo nelle colorazioni panottiche, risulta privo di
strutture o presenta rare granulazioni azzurrofile.
Al microscopio elettronico i piccoli linfociti presentano un nucleo con
cromatina in ammassi, con piccolo nucleolo frequentemente contornato da zolle di
eterocromatina, e circondato da una lamella di ergastoplasma delimitante uno
spazio perinucleare. Nel citoplasma si osservano corte lamelle
ergastoplasmatiche, con pochi ribosomi. Il corpo di Golgi è scarsamente
sviluppato e poco numerosi sono anche i lisosomi.
Per lungo tempo il piccolo linfocito è stato considerato elemento terminale non
più capace di divisioni mitotiche, destinato come tale ad esaurire le sue
funzioni; tuttavia i recenti dati di fisiopatologia hanno dimostrato la
possibilità che i linfociti fanno parte di un sistema complesso che ha capostipiti
nel midollo osseo e le cui fasi evo., comportano una serie di trasformazioni a
sec di stimoli appropriati e del microambiente di sviluppo.
 Dopo stimolazione antigenica il linfocito si trasforma in elementi di grandi
dimensioni (immunoblasti) che raggiungono 20-30 micron di diametro e posseggono
citoplasma fortemente basofilo, talora vacuolato. Il nucleo grande ed
arrotondato possiede cromatina reticolare e nucleoli ben visibili. Il passaggio
allo stadio di plasmacellula coincide con la formazione di elementi di foggia
ovalare con c metro di 12-20 micron lungo l'asse maggiore. Il nucleo è situato
in posizione eccentrica, con cromatina a zolle compatte simulanti, per
disposizione il dorso di una tartaruga. Il citoplasma è particolarmente
basofilo, con centrosoma chiaro in posizione paranucleare.
Dopo stimolazione antigenica il linfocito si trasforma in elementi di grandi
dimensioni (immunoblasti) che raggiungono 20-30 micron di diametro e posseggono
citoplasma fortemente basofilo, talora vacuolato. Il nucleo grande ed
arrotondato possiede cromatina reticolare e nucleoli ben visibili. Il passaggio
allo stadio di plasmacellula coincide con la formazione di elementi di foggia
ovalare con c metro di 12-20 micron lungo l'asse maggiore. Il nucleo è situato
in posizione eccentrica, con cromatina a zolle compatte simulanti, per
disposizione il dorso di una tartaruga. Il citoplasma è particolarmente
basofilo, con centrosoma chiaro in posizione paranucleare.
In condizioni fisiologiche, i linfociti sembrano una popolazione cellulare
omogenea. In realtà, dal punto di vista funzionale, negli elementi linfoidi sono
compresi i linfociti T, linfociti B e linfociti della «terza
sottopopolazione» (linfociti killer e natural killer) così denominati in
quanto non sicuramente riconducibili funzionalmente né ai linfociti T né ai B.
In verità i linfociti natural killer presentano una morfologia peculiare
rispetto agli elementi T e B, in quanto hanno dimensioni maggiori e citoplasma
più abbondante, ricco in granulazioni azzurrofile. Nell'ambito dei linfociti T
sono comprese almeno due sottopopolazioni morfologicamente indistinguibili: i
linfociti T helper (cooperatori) e linfociti T suppressor-cytotoxic
(soppressori).
Il linfocita T helper, identificato già a metà degli anni '70 per la presenza di
un recettore di membrana per la porzione Fc della immunoglobulina IgM (da cui
deriva la denominazione T) o per l'assenza dell'antigene TH2, specifico della
classe T suppressor (ed identificato dall'eteroantisiero di Reinherz ottenuto
da coniglio); viene oggi individuato per la positività all'anticorpo monoclonale
CD4.
Il linfocita «T suppressor-cytotoxic», identificato negli anni '70 per la
presenza della porzione Fc della immunoglobulina IgG (da cui il nome Ty) e per
la espressione dell'antigene di membrana TH2, risulta modernamente
caratterizzato da un corredo antigenico peculiare: positività nei confronti
degli anticorpi monoclonali di classe di differenziazione 8 (CD8), oltre che di
quelli caratteristici di tutti gli elementi linfocitari T maturi (CD3, CD2, CD5,
CD7). Svolge un'azione inibitoria nei confronti della risposta immunitaria B
cellulare, ed è in grado di esplicare un'attività citotossica nei confronti di
diverse classi cellulari o di sostanze microbiche.
I linfociti killer naturali (NK), morfologicamente sono distinguibili dalle
altre sottoclassi linfocitarie per le loro maggiori dimensioni cellulari (12-17
micron di diametro), per il nucleo inciso, l'ampia rima citoplasmatica, e la
presenza di granuli azzurrofili (da cui deriva la denominazione anglosassone «large
granular lymphocytes»). Sono così denominati per la loro capacità di agire in
assenza di una precedente sensibilizzazione e rappresentano una barriera
difensiva di prima linea nei confronti di aggressioni infettive e neoplastiche.
Esercitano una funzione citotossica diretta e spontanea, e sembrano svolgere un
ruolo importante nel rigetto dei trapianti midollari eterologhi, nella reazione GVH (graft versus host disease) e nella regolazione del processo maturativo
emopoietico midollare e timico attraverso una azione citolitica diretta, o
inibitoria sugli elementi staminali. Sono in grado di produrre linfochi-ne quali
gli interferoni, sostanze a spiccate attività antivirali e tumoricide, e
interleuchine.
I linfociti citotossici (killer, K), posseggono un'attività citotossica
cellulare anticorpo dipendente (ADCC) in quanto esercitano la loro funzione
immunitaria solamente su cellule rivestite da molecole anticorpali IgG, alle
quali si legano mediante un recettore specifico per il frammento Fc delle IgG
stesse. La conseguenza di questo legame è la lisi della cellula (da qui il nome
killer).

I linfociti B sono caratterizzati dalla presenza nella membrana di diversi antigeni di differenziazione. In particolare l'elemento maturo, reperibile nel sangue periferico, presenta i seguenti marcatori: recettori per le immunoglobuline le quali sono costantemente presenti sulla superficie cellulare; capacità di formare rosette EA (per la presenza del recettore per il frammento Fc delle IgG) e EAC (correlata alla presenza del recettore specifico per la frazione 3 del complemento: C3); espressione degli antigeni di classe II del MHC; positività nei confronti di anticorpi monoclonali di classe di differenziazione 19-20-21-22-23-24; specifici dei linfociti B e areattivi nei confronti degli elementi T.
Nell'organismo adulto, i linfociti sono prodotti e vanno incontro ad una prima
fase maturativa nel midollo osseo. Durante la vita embrionale, invece, altri
organi svolgono questa funzione: il sacco vitellino nei primi 3 mesi di
gestazione e successivamente il fegato e la milza. Le tappe differenziative che
contraddistinguono la maturazione dei linfociti T e B sono, però, totalmente
diversificate e un contributo sostanziale alla definizione delle stesse è
derivato dalla applicazione di tecniche citofluorimetriche e immunoenzimatiche
in associazione all'utilizzo di anticorpi monoclonali diretti contro antigeni
leucocitari di differenziazione ("cluster of designation"-CD). Queste strutture
(normalmente glicoproteine espresse sulla membrana citoplasmatica, o più
raramente nel compartimento citoplasmatico e/o nucleo) sono dotate di
specificità per la classe linfocitaria (T o B) o per una o più fasi di
maturazione dei linfociti T o B. Va comunque sottolineato il fatto che molti
anticorpi monoclonali che fino a qualche anno or sono erano considerati
specifici per alcuni citotipi ematici, in realtà presentano un pattern di
reattività complesso, reagendo anche con sottoclassi di altre classi cellulari.
Sia i linfociti B che T derivano da una comune cellula staminale ubicata nel
tessuto emopoietico midollare; le fasi successive di maturazione avvengono nel
midollo osseo (fase midollare di maturazione T e B linfocitaria), quindi a
livello timico per i linfociti T e ancora midollare per i linfociti B. Le fasi
conclusive dell'iter maturativo si compiono invece negli organi linfoidi
periferici (linfonodi, milza, tessuto linfatico dell'apparato digerente) ma in
regioni anatomicamente differenti per le due classi linfocitariei T e B.
- Linfopoiesi B. La maturazione dèi linfociti B si compie in due tappe
successive: la prima, di differenziazione centrale o midollare, denominata
"antigene indipendente",e la seconda, di differenziazione secondaria o
periferica, chiamata "antigene dipendente". Questa seconda fase maturativa si
svolge negli organi linfoidi periferici (milza, linfonodi, tessuto linfatico
dell'apparato digerente: tonsille, placche di Peyer, appendice, o del sistema
respiratorio e genito-urinario).
a) Maturazione antigene indipendente. I linfociti B derivano da una comune
cellula progenitrice staminale totipotente situata nel midollo osseo in grado di
originare elementi appartenenti a tutte le filiere cellulari ematiche. Da questo
elemento deriva una seconda cellula staminale commissionata in senso linfoide in
grado di originare probabilmente sia cellule indirizzate in senso T che B
linfocitario. Il successivo stadio maturativo coinvolge una cellula staminale
già orientata in senso B linfocitario. Numerosi marcatori fenotipici e
genotipici contrassegnano questa primitiva fase maturativa. Ci riferiamo
innanzitutto alla configurazione dei geni delle immunoglobuline che riarrangiano
in maniera specifica nei linfociti orientati in senso B linfoide, contrariamente
alle altre cellule ematopoietiche e non (ivi compresi i linfociti T) i cui geni
delle immunoglobuline rimangono in una configurazione
"germline" durante l'intero iter differenziativo. Dal punto di vista
morfologico, queste cellule hanno un aspetto blastico e si caratterizzano per la
presenza di alcuni marcatori quali l'enzima TdT (terminal desossinucleotidil
transferasi), una DNA polimerasi localizzata a livello nucleare, la quale può
essere identificata sia mediante tecniche immunologiche (attraverso l'uso di
anticorpi monoclonali) che biochimiche. Questi elementi definiti come "pro-B"
esprimono inoltre il CD19, CD72, antigeni di classe II del sistema maggiore di
istocompatibilità (HLA-DR), mentre non presentano immunoglobuline né in
superficie e né in citoplasma. In questa fase maturativa i linfociti esprimono
inoltre costante positività per l'antigene CD34, una proteina transmembrana
espressa in tutte le cellule progenitrici staminali del sistema emolinfopoietico.
La fase successiva di maturazione (denominata "common B") si caratterizza per la
presenza dell'antigene CD10 (noto anche come antigene CALLA, in quanto espresso
nella quasi totalità delle leucemie linfoblastiche acute -LAL- del bambino e nel
50% delle LAL dell'adulto). Altre molecole identificabili in questa fase
maturativa sono rappresentati dal CD19, CD72, CD34, TdT, HLA-DR, e il CD22
solamente a livello citoplasmatico (cCD22). La tappa successiva di maturazione
(denominata pre-B) è contraddistinta dalla comparsa di catene M
intracitoplasmatiche (clg), ma non dalla catena immunoglobulinica completa. La
reattività nei confronti dell'antigene CD10 e della TdT è nettamente diminuita o
talora totalmente assente. Positivi risultano il CD19, CD24, HLA-DR, CD72, CD24,
CD20 e CD22 citoplasmatico.
La fase successiva realizza la formazione del cosidetto linfocita "immaturo" o
"vergine", cosi chiamato in quanto ancora tollerante nei confronti degli stimoli
antigenici. Questo elemento può essere ritrovato sia nel midollo osseo che nel
sangue periferico o nei tessuti linfoidi periferici, e si caratterizza per la
presenza di una catena immunoglobulinica completa di tipo IgM sulla superficie
della cellula. Il linfocita B vergine può esprimere sia la catena leggera kappa
che lambda: nell'ontogenesi del linfocita B la sintesi di catene k precede
quella delle catene labda. Costante la positività per gli antigeni CD19, CD20,CD21
(recettore per il virus EBV),CD22 di membrana,CD24, CD72. Assenti il CD10
(CALLA), TdT e il CD34.
b) Maturazione antigene dipendente. Questa fase di sviluppo del linfocita B si
svolge negli organi linfoidi periferici i quali vengono raggiunti dai linfociti
midollari attraverso il circolo ematico dopo una fase di passaggio dal tessuto
emopoietico al sistema vascolare midollare. Le fasi successive di maturazione
variano a seconda dello stimolo antigenico che ne ha evocato la
differenziazione. Le tappe finali di trasformazione culminano, comunque, in
tutti i casi nella produzione di linfociti B altamente specializzati nella
risposta immunitaria quali le plasmacellule e i linfociti B memoria. Il
primo evento che si realizza in seguito a stimolazione antigenica è
rappresentato dalla produzione di immunoglobuline di tipo IgD, che vengono
assemblate sulla membrana plasmatica del B linfocita. I linfociti B IgD+
generalmente coesprimono, almeno in una prima fase,anche le IgM e rappresentano
una tappa essenziale nello sviluppo linfocitario B (denominato "linfocita
intermedio"), in quanto rendono il linfocita non più tollerante nei confronti
degli stimoli antigenici. Successivamente compaiono anche gli altri tipi di
catene pesanti (IgG e IgA) (fase di "linfocita B maturo") le quali in una prima
fase possono essere coespresse sulla superficie cellulare insieme alle catene
IgM e IgD, per poi essere perdute in una fase successiva di maturazione.
Esistono, però, sottoclassi linfocitarie B mature che mantengono per lunghi
periodi le catene pesanti IgM e IgD unitamente alle IgG o IgA. Caratteristica
dei linfociti B maturi è quella di presentare il fenomeno del "capping" ovvero
di raggruppare ad un polo della cellula le SIg; tale fenomeno è facilmente
svelabile al microscopio a fluorescenza in seguito ad applicazione di antisieri
anti-Ig.
L'ontogenesi del linfocita B si conclude con la trasformazione immunoblastica,
caratterizzata dalla espressione di SIg ad alta densità , CD10 e CD25 (recettore
per l'interleuchina 2), tappa che prelude alla formazione della plasmacellula
matura, la quale si contraddistingue per la presenza di notevoli quantità di
immunoglobuline all'interno del citoplasma (clg). Le plasmacellule sono inoltre
positive per il CD38, CD78, HLA-DR, mentre mancano dei tipici antigeni espressi
dai linfociti B maturi ed immaturi. Alcuni linfociti B maturi, in seguito ad
attivazione antigenica, assumono le caratteristiche di "linfocita B memoria",
così denominato in quanto rimane in uno stato di quiescenza in attesa di
riattivarsi in seguito al sopraggiungere di nuovo stimolo antigenico.
La maturazione dei linfociti T si compie in tre fasi
successive, e ciascuna delle quali si realizza in regioni anatomiche tra di loro
distinte. La prima fase maturativa avviene nel midollo osseo, organo che
provvede durante l'intero arco della vita ad un rifornimento continuo di cellule
staminali in grado di maturare in senso T. La seconda fase si compie nel timo,
organo che subisce una involuzione durante l'età adulta, il quale attraverso la
sintesi di ormoni specifici crea una situazione microambientale ideale per la
maturazione delle cellule T. I linfociti T, nella terza fase, migrano infine
negli organi linfatici periferici ove giungono a maturazione completa.
La caratterizzione immuno-fenotipica e bio-molecolare dei linfociti T ha dato un
contributo sostanziale nella definizione di questa composita ed eterogenea
classe cellulare. Ricordiamo che negli anni '70, ovvero prima dell'avvento
dell'era degli anticorpi monoclonali, solamente due marcatori identificavano la
classe linfocitaria T: la capacità di formare rosette con emazie di montone
(rosette E la reattività nei confronti dell'antisiero policlona HUTLA ("human T
lymphocyte antigen").
Fase midollare. E già stato ricordato come dal pool delle cellule staminali pluripotenti si sviluppino dapprima cellule progenitrici a duplice evolutività
in senso T e B e, quindi, cellule staminali oriertate in senso T. Il primo
elemento cellulare identificabile come appartenente alla serie linfocitaria T (fase pro-T) si caratterizza per la presenza dell'enzima TdT, e la espressione
di molecole quali HLA-DR, CD34 e talora CD38, CD7 e
cCD3 (CE : citoplasmatico).
Nessuna di queste molecole risulta però specifica per i linfociti T (ad
eccezione del cCD3). Per questo motivo il riconoscimento di linfociti T si deve
basare su tecniche bio-molecolari volte allo studio dei geni che sovraintendono
alla sintesi del "recettore per la cellula T" ("T-cell receptor" o TcR),
struttura specifica del linfocita T appartenente alla superfamiglia delle
immunoglobuline Attualmente sono disponibili anticorpi monoclonali con
reattività per il dimero alfa/beta del TcR dotati di una buona specificità. Sono
inoltre in commercio reagenti in grado di riconoscere la caten..-beta
citoplasmatica, la catena gamma o delta, o dimero gamma/ delta.
Fase timica. I linfociti midollari già orientati in senso T migrano nel timo,
colonizzando in un prime momento solamente le aree sottocapsulari. Questa fase,
denominata "pretimocita", o "pre-T" o di "timocita immaturo" o di "grande blasto
timico sottocapsulare", si caratterizza per la espressione dell'antigene CD7, glicoproteina di peso molecolare 40 KD espressa su tutti i linfociti timici, CD2
(recettore per gli eritrociti di montone responsabile della formazione delle
rosette E), CD38, TdT, e cCD3. I pre-timociti rappresentano il 3-5% dei timociti,
e si caratterizzano inoltre per la mancata reattività in vitro nei confronti di
sostanze mitogeniche (fitoemagglutinina -PHA, concanavalina A (Con-A)
caratteristicamente attive con i linfociti T maturi. Nonostante la positività
per l'antigene CD2, questi linfociti non sono ancora in grado di formare rosette
E. Ciò sembra dovuto al fatto che l'antigene CD2, in questa fase maturativa, è
ancora funzionalmente inattivo. E una fase di sviluppo, antigene indipendente,
che si contraddistingue per la elevata attività proliferativa. In una fase
successiva (timocita corticale o timocita comune) i timociti sottocapsulari
migrano nella regione corticale dove acquisiscono positività per il CDl, il
quale viene espresso solamente in questa fase differenziativa. I timociti
corticali esprimono inoltre il CD7, CD38, CD2, CD5 (molecola espressa anche nei
linfociti T maturi, e in una sottopopolazione di linfociti B) e coesprimono
nella stessa cellula il CD4 ed il CD8, le quali permet-tono il risconoscimento
delle due principali sottopo-lazioni linfocitarie T mature ("T helper" e "T
suppressor", rispettivamente). Circa il 70-80% della popolazione timica globale
è composta da timociti
comuni, gran parte dei quali (circa il 90%) va precocemente incontro a morte
nel contesto del timo e senza raggiungere mai una fase completa di maturazione
grazie al meccanismo di apoptosi regolato dalla proteina Bcl2.
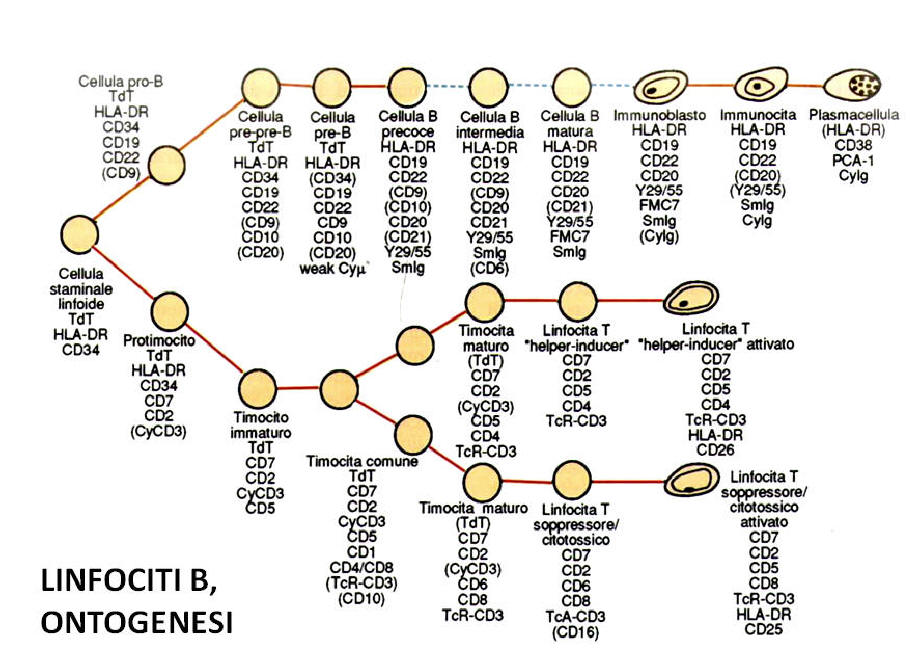
Fase periferica di maturazione con descrizione delle principali sottoclassi linfocitarie T.
In questa fase i linfociti T timici acquisiscono, sia dal punto
di vista funzionale che fenotipico, le caratteristiche proprie del linfocita T
maturo circolante, dal quale però se ne discostano per alcune caratteristiche
ridotta risposta mitogenica nei confronti del PHA e Con-A). Il "timocita
maturo", noto anche come
timocita midollare", può esprimere sia un fenotipo CD4+ (tipico della sottopolazione linfocitaria "T-helper-inducer") o
CD8+. Questi elementi si
caratterizzano inoltre per la positività per l'antigene CD3 di superficie,
struttura associata al "T-cell receptor", che media la trasduzione di diversi
segnali biochimici all'interno della cellula. Essendo la molecola CD3 parte del
TCR, il suo riconoscimento nei linfociti maturi riveste un ruolo diagnostico
essenziale e spesso insostituibile. L'antigene CD3 citoplasmatico, diversamente
da quello di superficie, è espresso, invece, anche nei timociti corticali e
talora sottocapsulari. Il timo svolge un ruolo chiave nello sviluppo dei
linfociti T, in quanto crea condizioni microambientali per la maturazione di
queste cellule. Le cellule epiteliali timiche producono numerosi ormoni (timosina,
timopoietina, fattore timico ormonale-THF, fattore timico sierico) che
interagiscono direttamente o indirettamente con recettori presenti sui linfociti
T avviati a maturazione. Fase ematica e periferica di sviluppo. Le fasi
successive di sviluppo si compiono negli argani linfoidi periferici in risposta
a stimoli antigenici di vario tipo. I linfociti T timici attraverso il circolo
ematico colonizzano aree specifiche nei linfonodi, milza, ecc. (cosidette aree "T-dipendenti",
ben separate da quelle B). Dopo contatto con antigeni i linfociti T acquisiscono
alcuni marcatori di superficie (denominati di attivazione) quali il ZD25
(recettore per IL2), CD26, CD38, HLA-DR, JD30 (linfociti attivati).
I linfociti CD4 + (linfociti T helper) del sangue periferico esistono in due
principali sottopopolazioni, tra loro distinguibili in base alla espressione di
alcuni marcatori immunologici: T helper/inducer CD4+, CD29+, CD45RO+), T suppressor/inducer
(CD4+, CD45RA+).
I linfociti CD4 + /CD45RA- identificano inoltre
linfociti T memoria, ovvero quella sottopopolaziole linfoide che ha già
incontrato l'antigene e che quindi risponderà in maniera più rapida e specifica ad un secondo o più contatto
antigenico.
I linfociti CD4+/CD45RA+ identificano i cosidetti linfociti "naive" (linfociti
T vergini che non hanno ancora avuto contatti con antigeni). Questa sottopopolazione linfocitaria T rappresenta il 90% lei linfociti T del cordone
ombelicale, per poi ridursi progressivamente nel bambino e assestarsi
nell'adulto attorno al 30-35%. Un'altro marcatore impiegabile per identificare i
linfociti "naive" sia di tipo B che T, è rappresentata dalla L-selectina
(molecola di adesione), la quale risulta espressa solamente nei linfociti non
stimolati (vergini).
I linfociti CD8+ possono svolgere una attività soppresssiva (CD8+/CD11b + ) o
citotossica (MHC ristretta: CD8+/CD11b-).
Esiste inoltre una sottoclasse di linfociti CD3+ la quale coesprime il CD57
(marcatore NK associato) che si caratterizza per l'attività citotossica non MHC
ristretta, del tutto sovrapponibile a quella dei linfociti NK (natural killer),
i quali caratterizzano la terza classe linfocitaria (non B non T) e che hanno
come attività funzionale specifica quella di esercitare una attività citolitica
non correlato ad un riconoscimento di antigeni del sistema HLA.
Le cellule NK hanno un fenotipo CD3—, CD2+, CD16+, CD56+, CD57+ / -. Questi
elementi rappresentano il 20% dei linfociti del sangue periferico del neonato e
si riducono progressivamente con l'età.
Il 95% dei linfociti T CD3+ del sangue periferico esprimono il dimero alfa-beta
del TCR, mentre solamente il 4% dei linfociti T CD3+ presentano positività per
il dimero gamma-delta del TCR.
Recentemente è stata messa a punto una nuova tecnica di biologia molecolare,
denominata TREC ("TCR-rearrangement excision circles"), che permette di
identificare tutti i linfociti di derivazione timica, denominati "recenti
emigranti timici". Il metodo è basato sul fatto che i linfociti T riarrangiano i
geni del TCR con lo scopo di produrre le proteine che formano il complesso
recettoriale TCR, che deve avere una grande variabilità per permettere il
riconoscimento di tutte le molecole antigeniche con le quali i linfociti possono
entrare in contatto. Il riarrangiamento implica l'eliminazione preventiva o
escissione di frammenti di DNA dal DNA genomico che assumono una forma
circolare. Tale frammenti, chiamati TREC ("TCR-rearrangement excision circles")
sono stabili, non vengono duplicati durante la mitosi, e contraddistinguono il
linfocita T che matura all'interno del timo. Sono in grado di offrire una stima
dei linfociti periferici di derivazione timica. Questo metodo offre enormi
opportunità in patologia e nel monitoraggio terapeutico in corso di tra-pianto
di cellule staminali periferiche, permettendo una accurata valutazione del danno
linfocitario T e di ripresa della funzione timica dopo trapianti.
Negli anni '90 sono state identificate almeno due nuove sottoclassi di linfociti
T helper, le quali giocano un ruolo chiave nei processi immunitari,
indirizzandoli nella risposta cellulo mediata in un caso o in quella umorale
nel secondo caso.
Le cellule TH1 mediano la risposta cellulomediata, fagocito-dipendente; possono
essere identificate per la loro capacità di produrre IL2, IFN-y, TNF-B,
anticorpi opsonizzanti o fissanti il complemento. Queste cellule stimolano la
proliferazione di linfociti T; dopo contatto antigenico attivano i macrofagi, la
citotossicità cellulare anticorpo dipendente e inducono resistenza alle
infezioni. I linfociti TH1 attivati esprimono i LAG-3 (lymphocyte activation
gene-3) che appartengono alla superfamiglia delle immunoglobuline. I TH1
esprimono inoltre 2 recettori per chemochine: CXCR-3 e CCR-5.
I linfociti TH2 mediano la risposta di tipo umorale fagocito-indipendente; si
caratterizzano per la capacità di produrre IL4, IL10, IL5, IL6 e IL13. Queste
cellule hanno una funzione inibente nei confronti dei macrofagi, stimolano la
sintesi di IgGl, IgG4, IgE, attivano le plasmacellule e gli eosinofili e sono
implicate nella suscettibilità alle infezioni. Contrariamente alle cellule TH1,
i linfociti TH2 attivati esprimo la molecola CD30, la quale fa parte della
superfamiglia dei recettori del TNF> oltre che alcuni recettori per chemochine:
CXCR4, CCR-3, CCR-4, CCR-7, e CCR-8.
Sia i linfociti TH1 che TH2 esistono in due principali sottopopolazioni di
linfociti CD4+: CD4 + a/b+ eCD4+ y/d+
Una terza sottopolazione di linfociti è denominata THO: presenta caratteristiche
intermedie tra i TH1 e TH2.
Esiste una altra sottopopolazione linfocitaria, denominata THp, che identifica i
linfociti T helper di tipo naive: questi non hanno ancora incontrato l'antigene
e vengono identificati con il termine p (precursori) in relazione alla loro
relativa immaturità rispetto ai linfociti T helper e soppressori maturi. Dopo
contatto con l'antigene ed in seguito ad interazioni con altri definenti del
sistema immunitario, queste cellule vanno incontro ad un processo di
differenziazione che culmina nella loro trasformazione in linfociti di tipo TH1,
TH2 o THO. In questa fase maturativa giocano un ruolo fondamentale le molecole
cosidette co-stimolatorie, che vengono secrete durante i processi di interazione
fra le cellule presentanti l'antigene (APC) e le cellule naive THp. Durante
questa fase, molecole del sistema HLA di classe II vengono presentate dalle
cellule APC sulla superfice dei linfociti THp,ed in particolare a livello del
complesso TCR. Questa interazione attiva la cellula THp che, come conseguenza,
esprime il recettore per la IL2, secerne IL2 e incrementa l'espressione del
ligando per il CD40. Queste modificazioni portano da un lato ad un aumento della
capacità del linfocita T di legare in maniera costitutiva la molecola CD40 sulla
superfice degli elementi APC; inoltre stimolano gli APC ad esprimere dapprima i
complessi CD86-B7-2 e quindi il complesso CD80-B7-1. Questi processi svolgono un
ruolo chiave nel differenziamento della cellula THp e nella acquisizione di
funzioni specifiche, a causa dei seguenti eventi: 1) incremento della secrezione
di IL2 con conseguente proliferazione linfocitaria 2) produzione di sostanze anti-
apoptotiche quali Bcl-x; 3) secrezione di citochine specifiche per maturazione
T-linfocitica.
La maturazione in senso TH2 è regolata in particolare dal sistema B7-CD28, ovvero
dal complesso CD86/B7-2, oltre che da ICOS, una molecola, quest'ultima,
costimulatoria di recente scoperta, e c una sostanza appartenente alla
superfamiglia TX F (tumor necrosis factor): OX40. Per quanto riguarda la
differenziazione in senso TH1, sembrano essere attive: la molecola 4-1BB e la
molecola CD80-B7-1attive, quest'ultima, anche nella maturazione TH2.