Notes by doctor. Claudio Italiano
Lymphocytes represent a heterogeneous population of leukocyte elements
distributed in various areas of the body. In the classical sense, they are made
up of small elements with poor cytoplasm, with a pienotic chromatin nucleus;
however, the term "lymphocytic cells" tends to refer to the entire complex of
elements that involves traditional lymphocytes (small, medium and large),
lymphoblasts and plasma cells as forms of activation of a definite category of
the same elements. Most peripheral blood lymphocytes are small 7-8 microns in
diameter (small lymphocytes), while a minor portion (10%) is 9-15 micron (medium
to large lymphocyte). The small lymphocytes appear to consist of a nucleus with
condensed, roundish or just indented cloned matrix, devoid of visible nucleoli,
occupying the whole cell except for a small strip of moderately basophilic
cytoplasm; this in the panoptic colors, is devoid of structures or presents rare
azure-like granulations. Under the electron microscope the small lymphocytes
have a nucleus with chromatin in clusters, with a small nucleol frequently
surrounded by plates of heterochromatin, and surrounded by a lamella of
ergastoplasma delimiting a perinuclear space. In the cytoplasm, ergastoplasmatic
lamellae are observed, with few ribosomes. The body of Golgi is poorly developed
and the lysosomes are few in number. For a long time the small lymphocyte was
considered a terminal element no longer capable of mitotic divisions, destined
as such to exhaust its functions; however, recent data from pathophysiology have
shown the possibility that lymphocytes are part of a complex system that has
progenitors in the bone marrow and whose evo phases, involve a series of
transformations according to appropriate stimuli and the microenvironment of
development. After antigenic stimulation the lymphocyte is transformed into
large elements (immunoblasts) that reach 20-30 microns in diameter and possess
strongly basophilic cytoplasm, sometimes vacuolated. The large and rounded
nucleus has reticular chromatin and clearly visible nucleoli. The transition to
the plasma cell stage coincides with the formation of oval shaped elements with
a diameter of 12-20 microns along the major axis. The nucleus is located in an
eccentric position, with compact simulated clods of chromatin, to lay the back
of a turtle. The cytoplasm is particularly basophilic, with a clear centrosome
in a paranuclear position. Under physiological conditions, lymphocytes appear to
be a homogeneous cell population. In fact, from the functional point of view,
the lymphocytes T, B lymphocytes and lymphocytes of the "third subpopulation"
(killer lymphocytes and natural killer) are so named in so far as they are
certainly not functionally traceable neither to T lymphocytes nor to B. In truth,
natural killer lymphocytes have a peculiar morphology with respect to the T and
B elements, as they have larger dimensions and more abundant cytoplasm, rich in
blue-white granulations. Within the T lymphocytes there are at least two
morphologically indistinguishable subpopulations: T helper lymphocytes (cooperators)
and suppressor-cytotoxic T lymphocytes (suppressors). T helper lymphocyte,
already identified in the mid-seventies due to the presence of a membrane
receptor for the Fc portion of the IgM immunoglobulin (from which the T name
derives) or for the absence of the TH2 antigen, specific to class T suppressor
(and identified by the Reinherz heteroanthedral obtained from rabbit); is now
identified for the positivity to the CD4 monoclonal antibody.
The T suppressor-cytotoxic lymphocyte, identified in the 1970s by the presence
of the Fc portion of the IgG immunoglobulin (hence the name Ty) and the
expression of the TH2 membrane antigen, is modernized by a peculiar antigenic
kit: positivity to monoclonal antibodies of differentiation class 8 (CD8), as
well as to those characteristic of all mature T lymphocyte elements (CD3, CD2,
CD5, CD7).
It
performs an inhibitory action against the cellular immune response B, and is
able to perform cytotoxic activity against different cellular classes or
microbial substances. The natural killer lymphocytes (NK), morphologically, are
distinguishable from the other lymphocyte subclasses for their larger cell size
(12-17 microns in diameter), for the incised nucleus, the wide cytoplasmic rhyme,
and the presence of azurophile granules (from which derives the Anglo-Saxon
denomination "large granular lymphocytes"). They are so named for their ability
to act in the absence of previous sensitization and represent a frontline
defensive barrier against infectious and neoplastic aggressions. They have a
direct and spontaneous cytotoxic function, and they seem to play an important
role in the rejection of heterologous bone marrow transplants, in the GVH
reaction (graft versus host disease) and in the regulation of the bone marrow
and thymol maturative maturation process through a direct cytolytic or
inhibitory action on the elements stem. They are capable of producing
lymphocytes such as interferons, substances with marked antiviral and
tumoricidal activity, and interleukins. Cytotoxic lymphocytes (killer, K) have a
cytotoxic antibody-dependent cellular activity (ADCC) as they exert their immune
function only on cells coated with IgG antibody molecules, to which they are
bound by a specific receptor for the IgG Fc fragment themselves. The consequence
of this link is the lysis of the cell (hence the name killer).
B lymphocytes are characterized by the presence in the membrane of different differentiation antigens. In particular, the mature element, available in the peripheral blood, has the following markers: receptors for immunoglobulins which are constantly present on the cell surface; ability to form EA rosettes (due to the presence of the Fc fragment of the IgG) and EAC (correlated to the presence of the specific receptor for the fraction 3 of the complement: C3); expression of class II antigens of the MHC; positivity to monoclonal antibodies of differentiation class 19-20-21-22-23-24; specific of B lymphocytes and areattivi in relation to the T elements.
In the adult organism, the lymphocytes are produced and undergo a first stage
of maturation in the bone marrow. During embryonic life, however, other organs
perform this function: the yolk sac in the first 3 months of gestation and then
the liver and spleen. The differentiation stages that distinguish the maturation
of T and B lymphocytes are, however, totally diversified and a substantial
contribution to their definition is derived from the application of
cytofluorimetric and immunoenzymatic techniques in association with the use of
monoclonal antibodies directed against differentiation leukocyte antigens (
"cluster of designation" -CD). These structures (normally glycoproteins
expressed on the cytoplasmic membrane, or more rarely in the cytoplasmic and /
or nucleus compartment) are specific for the lymphocyte class (T or B) or for
one or more maturation phases of the T or B lymphocytes. underlined the fact
that many monoclonal antibodies that up until a few years ago were considered
specific for some blood cytotypes, actually present a pattern of complex
reactivity, also reacting with subclasses of other cell classes.
Both B and T lymphocytes derive from a common stem cell located in the medullary
hemopoietic tissue; the successive stages of maturation take place in the bone
marrow (medullary phase of T and B lymphocyte maturation), then at thymic level
for T lymphocytes and again medullary for B lymphocytes. The final stages of
maturation are performed in the peripheral lymphoid organs (lymph nodes, spleen,
lymphatic system of the digestive tract) but in anatomically different regions
for the two lymphocytic classes T and B.
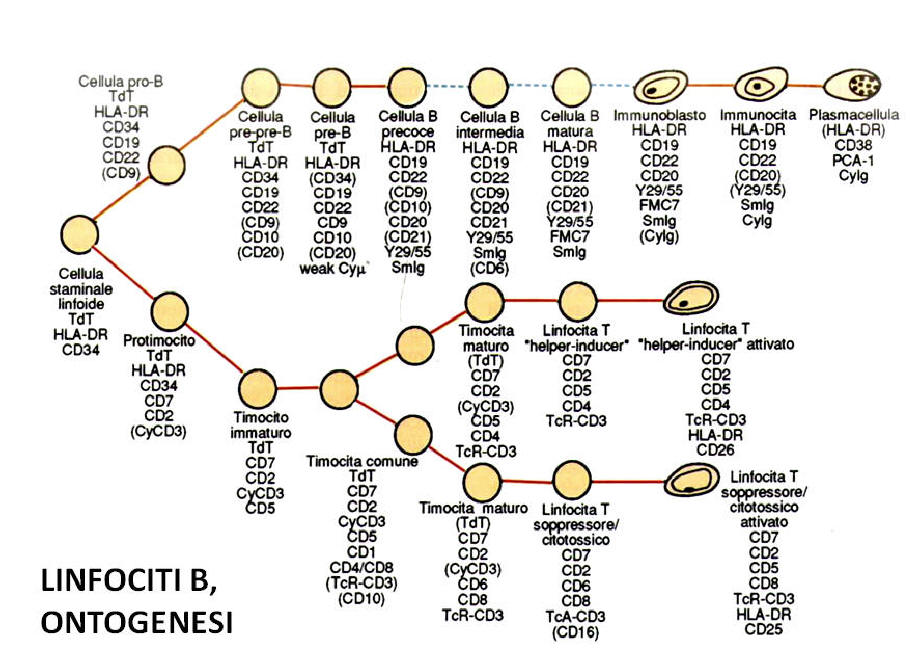
- B lymphopoiesis. The maturation of B lymphocytes takes place in two successive
stages: the first, of central or medullary differentiation, called "independent
antigen", and the second, of secondary or peripheral differentiation, called "dependent
antigen". This second stage of maturation takes place in the peripheral lymphoid
organs (spleen, lymph nodes, lymphatic tissue of the digestive system: tonsils,
Peyer's plaques, appendix, or of the respiratory and genitourinary systems).
a) Independent antigen maturation. B lymphocytes derive from a common totipotent
stem progenitor cell located in the bone marrow capable of originating elements
belonging to all the blood cell lines. From this element derives a second stem
cell commissioned in the lymphoid direction able to probably originate both
T-cell and B-lymphocyte-oriented cells. The subsequent maturation stage involves
a stem cell already oriented in the B-lymphocytic direction. Numerous phenotypic
and genotypic markers mark this primitive maturation phase. We refer above all
to the configuration of the immunoglobulin genes that specifically rearrange in
lymphocyte-oriented lymphocytes, in contrast to other hematopoietic and
non-hematopoietic cells (including T cells) whose immunoglobulin genes remain in
a "germline" configuration during 'whole differentiation process. From the
morphological point of view, these cells have a blastic appearance and are
characterized by the presence of some markers such as the TdT (terminal
deoxynucleotidyl transferase) enzyme, a DNA polymerase located at the nuclear
level, which can be identified by immunological techniques ( through the use of
monoclonal antibodies) that biochemical. These elements defined as "pro-B" also
express CD19, CD72, class II antigens of the major histocompatibility system (HLA-DR),
while they do not exhibit immunoglobulins neither on the surface nor in
cytoplasm. In this maturation phase the lymphocytes also express a constant
positivity for the CD34 antigen, a transmembrane protein expressed in all the
stem progenitor cells of the hemolymphopoietic system.
The next phase of maturation (called "common B") is characterized by the
presence of the CD10 antigen (also known as CALLA antigen, as expressed in
almost all acute lymphoblastic leukemia -LAL- of the child and in 50% of the LAL
'adult). Other identifiable molecules in this maturation phase are CD19, CD72,
CD34, TdT, HLA-DR, and CD22 only at the cytoplasmic level (cCD22). The next
stage of maturation (called pre-B) is marked by the appearance of
intracytoplasmic M chains (clg), but not by the complete immunoglobulin chain.
The reactivity towards the CD10 antigen and the TdT is clearly decreased or
sometimes totally absent. CD19, CD24, HLA-DR, CD72, CD24, CD20 and cytoplasmic
CD22 are positive.
The next phase realizes the formation of the so-called "immature" or "virgin"
lymphocyte, so called because it is still tolerant towards antigenic stimuli.
This element can be found either in the bone marrow or in the peripheral blood
or in the peripheral lymphoid tissues, and is characterized by the presence of a
complete immunoglobulin chain of the IgM type on the cell surface. The virgin B
lymphocyte can express both the light chain kappa and lambda: in the ontogenesis
of the B lymphocyte the synthesis of k chains precedes that of the labda chains.
Positive for the antigens CD19, CD20, CD21 (receptor for EBV virus), membrane
CD22, CD24, CD72. Absent CD10 (CALLA), TdT and CD34.
b) Dependent antigen maturation. This development phase of the B lymphocyte
takes place in the peripheral lymphoid organs which are reached by the medullary
lymphocytes through the blood circulation after a phase of transition from the
haematopoietic tissue to the medullary vascular system. The successive stages of
maturation vary according to the antigenic stimulus that has evoked
differentiation. The final stages of transformation culminate, however, in all
cases in the production of B lymphocytes highly specialized in the immune
response such as plasma cells and memory B lymphocytes. The first event that
occurs following antigenic stimulation is represented by the production of IgD
immunoglobulins, which are assembled on the plasma membrane of the B lymphocyte.
B IgD + lymphocytes generally coexpress, at least in a first phase, also the IgM
and represent an essential stage in the B lymphocyte development (called
"intermediate lymphocyte"), as they make the lymphocyte no more tolerant towards
antigenic stimuli. Then the other types of heavy chains (IgG and IgA) ("mature B
lymphocyte" phase) appear, which in a first phase can be coexpressed on the cell
surface together with the IgM and IgD chains, and then lost in a subsequent
phase of maturation. However, there are mature B lymphocyte subclasses that
maintain the IgM and IgD heavy chains together with IgG or IgA for long periods.
Characteristic of mature B lymphocytes is to present the phenomenon of "capping"
or to group the SIGs at a pole of the cell; this phenomenon can easily be
detected under a fluorescence microscope following application of anti-Ig
antisera.
The ontogenesis of the B lymphocyte ends with the immunoblastic transformation,
characterized by the expression of high-density SIg, CD10 and CD25 (receptor for
interleukin 2), a step that is a prelude to the formation of the mature plasma
cell, which is characterized by the presence of significant amounts of
immunoglobulins within the cytoplasm (clg). The plasma cells are also positive
for CD38, CD78, HLA-DR, while they lack the typical antigens expressed by mature
and immature B lymphocytes. Some mature B lymphocytes, following antigenic
activation, assume the characteristics of "memory B lymphocyte", so called
because it remains in a state of quiescence waiting to be reactivated following
the arrival of a new antigenic stimulus.
The maturation of T lymphocytes takes place in three successive phases, each
of which is carried out in separate anatomical regions. The first stage of
maturation takes place in the bone marrow, an organ that provides throughout the
entire life span with a continuous supply of stem cells able to mature in the T
direction. The second phase is performed in the thymus, an organ that undergoes
an involution during the adulthood, which through the synthesis of specific
hormones creates an ideal micro-environmental situation for the maturation of T
cells. The T lymphocytes, in the third phase, migrate finally in the peripheral
lymphatic organs where they reach full maturation.
The immuno-phenotypic and bio-molecular characterization of T lymphocytes has
made a substantial contribution in defining this composite and heterogeneous
cell class. Recall that in the '70s, ie before the advent of the monoclonal
antibody era, only two markers identified the T lymphocyte class: the ability to
form rosettes with ram's blood cells (rosettes AND the reactivity towards the
polyclonal antiserum HUTLA ( "human T lymphocyte antigen").
Medullary phase. It has already been recalled that from the pool of pluripotent
stem cells first develop progenitor cells with a dual evolution in the T and B
direction and, therefore, stem cells oriented in T direction. The first cellular
element identifiable as belonging to the T lymphocyte series (pro- T) is
characterized by the presence of the TdT enzyme, and the expression of molecules
such as HLA-DR, CD34 and sometimes CD38, CD7 and cCD3 (CE: cytoplasmic). However,
none of these molecules is specific for T lymphocytes (with the exception of
cCD3). For this reason the recognition of T lymphocytes must be based on
bio-molecular techniques aimed at the study of genes that supervise the
synthesis of the "T cell receptor" ("T-cell receptor" or TcR), specific
structure of the T lymphocyte belonging to to the immunoglobulin superfamily
Currently monoclonal antibodies with reactivity for the TcR alpha / beta dimer
are available with a good specificity. In addition, reagents capable of
recognizing the cytoplasmic beta-beta, the gamma or delta chain, or gamma /
delta dimer are commercially available.
Timothy phase. The T-oriented medullary lymphocytes migrate in the thymus,
colonizing in the first moment only the subcapsular areas. This phase, called "pretimocyte",
or "pre-T" or "immature thymocyte" or "large subcapsular thymic blast", is
characterized by the expression of the CD7 antigen, glycoprotein of 40 KD
molecular weight expressed on all lymphocytes thymomas, CD2 (receptor for sheep
erythrocytes responsible for the formation of rosettes E), CD38, TdT, and cCD3.
The pre-thymocytes represent 3-5% of thymocytes, and are also characterized by
the lack of in vitro reactivity against mitogenic substances (phytohaemagglutinin-PHA,
concanavalin A (Con-A) characteristically active with mature T lymphocytes.
positivity for the CD2 antigen, these lymphocytes are not yet able to form
rosettes E. This seems to be due to the fact that the CD2 antigen, in this
maturation phase, is still functionally inactive and is an independent antigen
development phase that It is characterized by high proliferative activity.In a
subsequent phase (cortical thymocyte or common thymocyte) subcapsular thymocytes
migrate in the cortical region where they acquire positivity for the CDl, which
is expressed only in this differentiation phase.The cortical thymocytes also
express the CD7 , CD38, CD2, CD5 (molecule expressed also in mature T
lymphocytes, and in a subpopulation of B lymphocytes) and coexpress in the same
cellulose a CD4 and CD8, which allow the recognition of the two main mature T
lymphocyte subpopulations ("T helper" and "T suppressor", respectively). About
70-80% of the global thymic population is composed of common thymocytes, most of
which (about 90%) precociously go to death in the context of the thymus and
without ever achieving a complete phase of maturation thanks to the regulated
apoptosis mechanism from the Bcl2 protein.
Peripheral maturation phase with description of the main T lymphocyte
subclasses
In this phase T-cell lymphocytes acquire, both from a functional and phenotypic
point of view, the characteristics typical of the circulating mature T
lymphocyte, from which however they depart for some characteristics reduced
mitogenic response to PHA and Con-A). The "mature thymocyte", also known as
medullary thymocyte ", can express either a CD4 + phenotype (typical of the"
T-helper-inducer "lymphocyte subpopulation) or CD8 +.These elements are also
characterized by the positivity for the surface CD3 antigen. , structure
associated to the "T-cell receptor", which mediates the transduction of
different biochemical signals inside the cell.That is the CD3 molecule part of
the TCR, its recognition in mature lymphocytes plays an essential and often
irreplaceable diagnostic role. Cytoplasmic CD3 antigen, unlike the surface
antigen, is expressed, however, also in cortical and sometimes subcapsular
thymocytes.The thymus plays a key role in the development of T lymphocytes, as
it creates micro-environmental conditions for the maturation of these cells.
thymic products produce numerous hormones (thymosin, thymopoietin, thymic
hormonal factor-THF, serum thymic factor) that interact directly or indirectly
with receptors present on T lymphocytes initiated when ripe. Blood phase and
peripheral development. The successive stages of development are accomplished in
peripheral lymphoid arganis in response to antigenic stimuli of various types. T
lymphocytes through the bloodstream colonize specific areas in the lymph nodes,
spleen, etc. (so-called "T-dependent" areas, well separated from those B). After
contact with antigens T lymphocytes acquire some surface markers (called
activation cells) such as ZD25 (IL2 receptor), CD26, CD38, HLA-DR, JD30 (activated
lymphocytes).
The CD4 + lymphocytes (helper T lymphocytes) of the peripheral blood exist in
two main subpopulations, distinguished among them by the expression of some
immunological markers: T helper / inducer CD4 +, CD29 +, CD45RO +), T suppressor
/ inducer (CD4 +, CD45RA +) .
The CD4 + / CD45RA-lymphocytes also identify T memory cells, that is, the
lymphoid subpoloid that has already encountered the antigen and will therefore
respond more quickly and specifically to a second or more antigenic contact.
The CD4 + / CD45RA + lymphocytes identify so-called "naive" lymphocytes (virgin
T lymphocytes that have not yet been in contact with antigens). This lymphocyte
subpopulation T represents 90% lei T lymphocytes of the umbilical cord, and then
gradually decreases in the child and settles in the adult around 30-35%. Another
marker that can be used to identify "naive" lymphocytes of both type B and T, is
represented by L-selectin (adhesion molecule), which is expressed only in
unstimulated (virgin) lymphocytes.
CD8 + lymphocytes may carry out a suppressive activity (CD8 + / CD11b +) or
cytotoxic (restricted MHC: CD8 + / CD11b-).
There is also a subclass of CD3 + lymphocytes which co-enacts CD57 (associated
NK marker) which is characterized by the non-MHC restricted cytotoxic activity,
completely overlapping that of NK (natural killer) lymphocytes, which
characterize the third lymphocyte class ( non B not T) and that have a specific
functional activity to exercise a cytolytic activity not related to an antigen
recognition of the HLA system. NK cells have a CD3-, CD2 +, CD16 +, CD56 +, CD57
+ / - phenotype. These elements represent 20% of the peripheral blood
lymphocytes of the newborn and are progressively reduced with age.
The 95% of the peripheral blood CD3 + T lymphocytes express the TCR
alpha-beta dimer, while only 4% of the CD3 + T lymphocytes have a positive
effect on the TCR gamma-delta dimer.
Recently a new molecular biology technique has been developed, called TREC ("TCR-rearrangement
excision circles"), which allows to identify all the thymocytes of thymic
derivation, called "recent timid emigrants". The method is based on the fact
that T lymphocytes rearrange the TCR genes with the aim of producing the
proteins that form the TCR receptor complex, which must have a great variability
to allow the recognition of all the antigenic molecules with which the
lymphocytes can get in touch. The rearrangement involves the preventive
elimination or excision of DNA fragments from the genomic DNA that take on a
circular shape. These fragments, called TREC ("TCR-rearrangement excision
circles") are stable, are not duplicated during mitosis, and distinguish the T
lymphocyte that matures within the thymus. They are able to offer an estimate of
peripheral lymphocytes of thymic derivation. This method offers enormous
opportunities in pathology and in the ongoing therapeutic monitoring of
peripheral stem cells, allowing an accurate evaluation of the T cell lymphocyte
damage and recovery of the thymic function after transplantation.
In the 1990s, at least two new subclasses of T helper lymphocytes have been
identified, which play a key role in immune processes, directing them in the
mediated cell response in one case or in the humoral one in the second case.
TH1 cells mediate the cell-mediated, phagocyte-dependent response; they can be
identified for their ability to produce IL2, IFN-y, TNF-B, opsonizing or
complement-fixing antibodies. These cells stimulate the proliferation of T
lymphocytes; after antigenic contact activate macrophages, cell cytotoxicity,
antibody dependent and induce resistance to infections. Activated TH1
lymphocytes express LAG-3 (lymphocyte activation gene-3) belonging to the
immunoglobulin superfamily. TH1 also express 2 chemokine receptors: CXCR-3 and
CCR-5.
TH2 lymphocytes mediate the phagocyte-independent humoral type response; they
are characterized by their ability to produce IL4, IL10, IL5, IL6 and IL13.
These cells have an inhibitory function against macrophages, stimulate the
synthesis of IgG1, IgG4, IgE, activate plasma cells and eosinophils and are
implicated in susceptibility to infections. Contrary to TH1 cells, activated TH2
lymphocytes express the CD30 molecule, which is part of the TNF receptor
superfamily> as well as some chemokine receptors: CXCR4, CCR-3, CCR-4, CCR-7,
and CCR-8 .
Both the TH1 and TH2 lymphocytes exist in two main subpopulations of CD4 +
lymphocytes: CD4 + a / b + eCD4 + y / d +
A third subpopulation of lymphocytes is called THO: it has intermediate
characteristics between TH1 and TH2.
There is another lymphocyte subpopulation, called THp, which identifies naive
helper T cells: these have not yet encountered the antigen and are identified by
the term p (precursors) in relation to their relative immaturity with respect to
the helper T lymphocytes and suppressors mature. After contact with the antigen
and following interactions with other immune system definers, these cells
undergo a differentiation process that culminates in their transformation into
TH1, TH2 or THO type lymphocytes. In this maturation phase, the so-called
co-stimulatory molecules play a fundamental role, which are secreted during the
interaction processes between the antigen-presenting cells (APC) and the
THp-naive cells. During this phase, molecules of the class II HLA system are
presented by the APC cells on the surface of the THp lymphocytes, and in
particular at the TCR complex level. This interaction activates the THp cell
which, as a consequence, expresses the receptor for IL2, secretes IL2 and
increases the expression of the ligand for CD40. These modifications lead on the
one hand to an increase in the T lymphocyte capacity of constitutively binding
the CD40 molecule on the surface of the APC elements; they also stimulate APCs
to first express the CD86-B7-2 complexes and then the CD80-B7-1 complex. These
processes play a key role in the differentiation of the THp cell and in the
acquisition of specific functions, due to the following events: 1) increase of
IL2 secretion with consequent lymphocytic proliferation 2) production of
anti-apoptotic substances such as Bcl-x; 3) secretion of specific cytokines for
T-lymphocytic maturation.
The maturation in the TH2 direction is regulated in particular by the B7-CD28
system, that is the CD86 / B7-2 complex, as well as ICOS, a molecule, the latter,
recently discovered costimulatory, and a substance belonging to the superfamily
TX F ( tumor necrosis factor): OX40. As far as TH1 differentiation is concerned,
the 4-1BB molecule and the CD80-B7-1active molecule, the latter, also appear in
TH2 maturation.
Hematology
{kind=link}